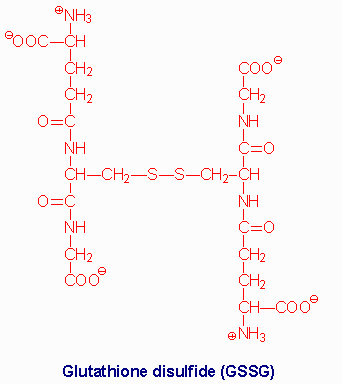Norepinephrine is the principal neurotransmitter of sympathetic postganglionic endings. Both norepinephrine and the methylated derivative, epinephrine are stored in synaptic knobs of neurons that secrete it, however, epinephrine is not a mediator at postganglionic sympathetic endings.
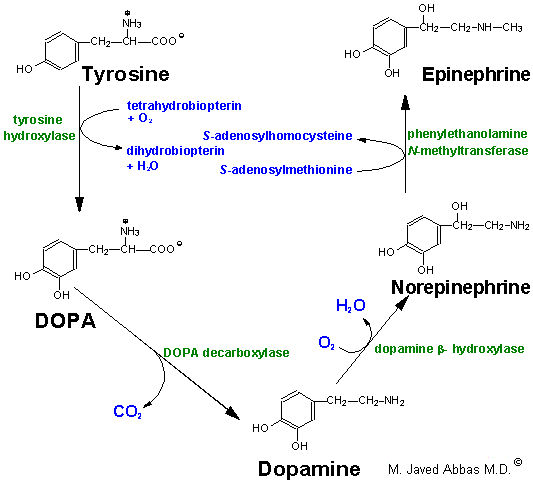
Tyrosine is transported into catecholamine-secreting neurons and adrenal medullary cells where catechaolamine synthesis takes place. The first step in the process requires tyrosine hydroxylase, which like phenylalanine hydroxylase requires tetrahydrobiopterin as cofactor. The hydroxylation reaction generates DOPA (3,4-dihydrophenylalanine). DOPA decarboxylase converts DOPA to dopamine, dopamine b-hydroxylase converts dopamine to norepinephrine and phenylethanolamine N-methyltransferase converts norepinephrine to epinephrine. This latter reaction is one of several in the body that uses SAM as a methyl donor generating S-adenosylhomocysteine. Within the substantia nigra and some other regions of the brain, synthesis proceeds only to dopamine. Within the adrenal medulla dopamine is converted to norepinephrine and epinephrine.
Synthesis of the catecholamines from tyrosine.
Once synthesized, dopamine, norepinephrine and epinephrine are packaged in granulated vesicles. Within these vesicles, norepinephrine and epinephrine are bound to ATP and a protein called chromogranin A.
Metabolism of the catecholemines occurs through the actions of catecholamine-O-methyltransferase, (COMT) and monoamine oxidase, (MAO). Both of these enzymes are widley distributed throughout the body. However, COMT is not found in nerve endings as is MAO.
back to the top
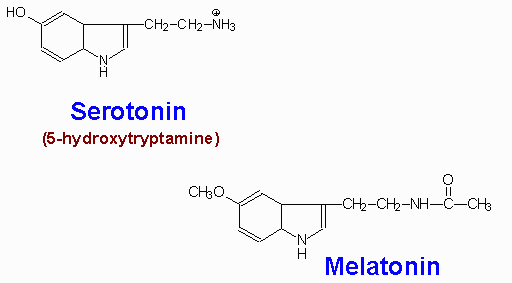
Serotonin is synthesized through 2-step process involving a tetrahydrobiopterin-dependent hydroxylation reaction (catalyzed by tryptophan-5-monooxygenase) and then a decarboxylation catalyzed by aromatic L-amino acid decarboxylase. The hydroxylase is normally not saturated and as a result, an increased uptake of tryptophan in the diet will lead to increased brain serotonin content.
Serotonin is present at highest concentrations in platelets and in the gastrointestinal tract. Lesser amounts are found in the brain and the retina. Serotonin containing neurons have their cell bodies in the midline raphe nuclei of the brain stem and project to portions of the hypothalamus, the limbic system, the neocortex and the spinal cord. After release from serotonergic neurons, most of the released serotonin is recaptured by an active reuptake mechanism. The function of the antidepressant, Prozac is to inhibit this reuptake process, thereby, resulting in prolonged serotonin presence in the synaptic cleft.
The function of serotonin is exerted upon its interaction with specific receptors. Several serotonin receptors have been cloned and are identified as 5HT1, 5HT2, 5HT3, 5HT4, 5HT5, 5HT6, and 5HT7. Within the 5HT1 group there are subtypes 5HT1A, 5HT1B, 5HT1D, 5HT1E, and 5HT1F. There are three 5HT2 subtypes, 5HT2A, 5HT2B, and 5HT2C as well as two 5HT5 subtypes, 5HT5a and 5HT5B. Most of these receptors are coupled to G-proteins that affect the activities of either adenylate cyclase or phospholipase Cg (PLCg). The 5HT3 class of receptors are ion channels.
Some serotonin receptors are presynaptic and others postsynaptic. The 5HT2A receptors mediate platelet aggregation and smooth muscle contraction. The 5HT2C receptors are suspected in control of food intake as mice lacking this gene become obese from increased food intake and are also subject to fatal seizures. The 5HT3 receptors are present in the gastrointestinal tract and are related to vomiting. Also present in the gastrointestinal tract are 5HT4 receptors where they function in secretion and peristalsis. The 5HT6 and 5HT7 receptors are distributed throughout the limbic system of the brain and the 5HT6 receptors have high affinity for antidepressant drugs.
Melatonin is derived from serotonin within the pineal gland and the retina, where the necessary N-acetyltransferase enzyme is found. The pineal parenchymal cells secrete melatonin into the blood and cerebrospinal fluid. Synthesis and secretion of melatonin increases during the dark period of the day and is maintained at a low level during daylight hours. This diurnal variation in melatonin synthesis is brought about by norepinephrine secreted by the postganglionic sympathetic nerves that innervate the pineal gland. The effects of norepinephrine are exerted through interaction with b-adrenergic receptors. This leads to increased levels of cAMP, which in turn activate the N-acetyltransferase required for melatonin synthesis. Melatonin functions by inhibiting the synthesis and secretion of other neurotransmitters such as dopamine and GABA.
back to the top
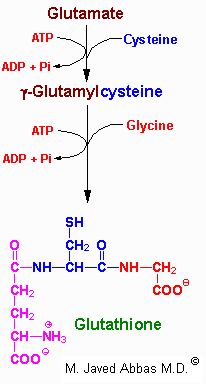
Several mechanisms exist for the transport of amino acids across cell membranes. Many are symport or antiport mechanisms that couple amino acid transport to sodium transport. The g-glutamyl cycle is an example of a group transfer mechanism of amino acid transport. Although this mechanism requires more energy input, it is rapid and has a high capacity. The cycle functions primarily in the kidney, particularly renal epithelial cells. The enzyme g-glutamyl transpeptidase is located in the cell membrane and shuttles GSH to the cell surface to interact with an amino acid. Reaction with an amino acid liberates cysteinylglycine and generates a g-glutamyl-amino acid which is transported into the cell and hydrolyzed to release the amino acid. Glutamate is released as 5-oxoproline and the cysteinylglycine is cleaved to its component amino acids. Regeneration of GSH requires an ATP-dependent conversion of 5-oxoproline to glutamate and then the 2 additional moles of ATP that are required during the normal generation of GSH.
back to the top
NOS is a very complex enzyme, employing five redox cofactors: NADPH, FAD, FMN, heme and tetrahydrobiopterin. NO can also be formed from nitrite, derived from vasodilators such as glycerin trinitrate during their metabolism. The half-life of NO is extremely short, lasting only 2-4 seconds. This is because it is a highly reactive free radical and interacts with oxygen and superoxide. NO is inhibited by hemoglobin and other heme proteins which bind it tightly.
Chemical inhibitors of NOS are available and can markedly decrease production of NO. The effect is a dramatic increase in blood pressure due to vasoconstriction. Another important cardiovascular effect of NO is exerted through the production of cGMP, which acts to inhibit platelet aggregation.
back to the top Back to Topics<<<<
This article has been modified by Dr. M. Javed Abbas. If you have any comments please do not hesitate to sign my Guest Book.
20:48 21/12/2002