Transcription is the mechanism by which a template strand of DNA is utilized by specific RNA polymerases to generate one of the three different classifications of RNA. These 3 RNA classes are:
1. Messenger RNAs (mRNAs): This class of RNAs are the genetic coding templates used by the translational machinery to determine the order of amino acids incorporated into an elongating polypeptide in the process of translation.
2. Transfer RNAs (tRNAs): This class of small RNAs form covalent attachments to individual amino acids and recognize the encoded sequences of the mRNAs to allow correct insertion of amino acids into the elongating polypeptide chain.
3. Ribosomal RNAs (rRNAs): This class of RNAs are assembled, together with numerous ribosomal proteins, to form the ribosomes. Ribosomes engage the mRNAs and form a catalytic domain into which the tRNAs enter with their attached amino acids. The proteins of the ribosomes catalyze all of the functions of polypeptide synthesis.
All RNA polymerases are dependent upon a DNA template in order to synthesize RNA. The resultant RNA is, therefore, complimentary to the template strand of the DNA duplex and identical to the non-template strand. The non-template strand is called the coding strand because its' sequences are identical to those of the mRNA. However, in RNA, U is substituted for T.
back to the top
E. coli RNA polymerase is composed of 5 distinct polypeptide chains. Association of several of these generates the RNA polymerase holoenzyme. The sigma subunit is only transiently associated with the holoenzyme. This subunit is required for accurate initiation of transcription by providing polymerase with the proper cues that a start site has been encountered.
In both prokaryotic and eukaryotic transcription the first incorporated ribonucleotide is a purine and it is incorporated as a triphosphate. In E. coli several additional nucleotides are added before the sigma subunit dissociates.
Elongation involves the addition of the 5'-phosphate of ribonucleotides to the 3'-OH of the elongating RNA with the concomitant release of pyrophosphate. Nucleotide addition continues until specific termination signals are encountered. Following termination the core polymerase dissociates from the template. The core and sigma subunit can then reassociate forming the holoenzyme again ready to initiate another round of transcription.
In E. coli transcriptional termination occurs by both factor-dependent and factor-independent means. Two structural features of all E. coli factor-independently terminating genes have been identified. One feature is the presence of 2 symmetrical GC-rich segments that are capable of forming a stem-loop structure in the RNA and the second is a downstream A rich sequence in the template. The formation of the stem-loop in the RNA destabilizes the association between polymerase and the DNA template. This is further destabilized by the weaker nature of the AU base pairs that are formed, between the template and the RNA, following the stem-loop. This leads to dissociation of polymerase and termination of transcription. Most genes in E. coli terminate by this method.
Factor-dependent termination requires the recognition of termination sequences by the termination protein, rho. The rho factor recognizes and binds to sequences in the 3' portion of the RNA. This binding destabilizes the polymerase-template interaction leading to dissociation of the polymerase and termination of transcription.
back to the top
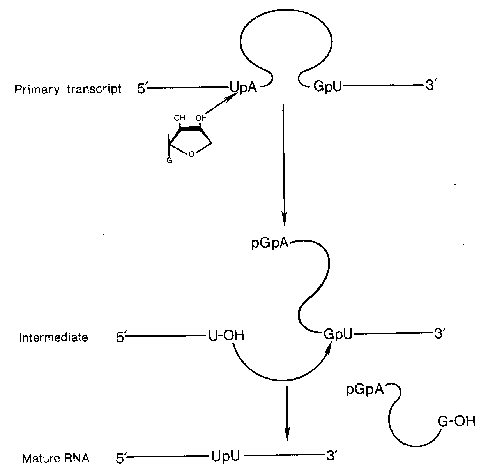
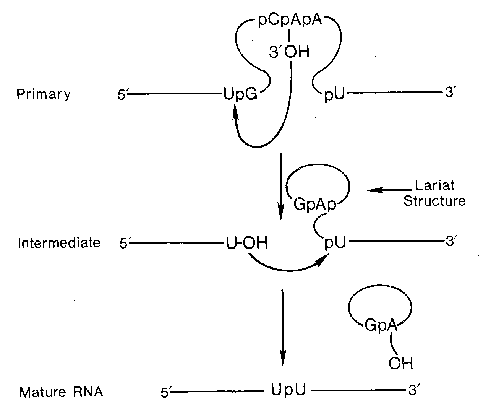
The presence of introns in eukaryotic genes would appear to be an extreme waste of cellular energy when considering the number of nucleotides incorporated into the primary transcript only to be removed later as well as the energy utilized in the synthesis of the splicing machinery. However, the presence of introns can protect the genetic makeup of an organism from genetic damage by outside influences such as chemical or radiation. An additionally important function of introns is to allow alternative splicing to occur, thereby, increasing the genetic diversity of the genome without increasing the overall number of genes. By altering the pattern of exons, from a single primary transcript, that are spliced together different proteins can arise from the processed mRNA from a single gene. Alternative splicing can occur either at specific developmental stages or in different cell types.
This process of alternative splicing has been identified to occur in the primary transcripts from at least 40 different genes. Depending upon the site of transcription, the calcitonin gene yields an RNA that synthesizes calcitonin (thyroid) or calcitonin-gene related peptide (CGRP, brain). Even more complex is the alternative splicing that occurs in the a-tropomyosin transcript. At least 8 different alternatively spliced a-tropomyosin mRNAs have been identified.
Abnormalities in the splicing process can lead to various disease states. Many defects in the b-globin genes are known to exist leading to b-thalassemias. Some of these defects are caused by mutations in the sequences of the gene required for intron recognition and, therefore, result in abnormal processing of the b-globin primary transcript.
Patients suffering from a number of different connective tissue diseases exhibit humoral auto-antibodies that recognize cellular RNA-protein complexes. Patients suffering from systemic lupus erythematosis have auto-antibodies that recognize the U1 RNA of the spliceosome.
back to the top Back to Topics<<<<
This article has been modified by Dr. M. Javed Abbas. If you have any comments please do not hesitate to sign my Guest Book.
20:49 21/12/2002