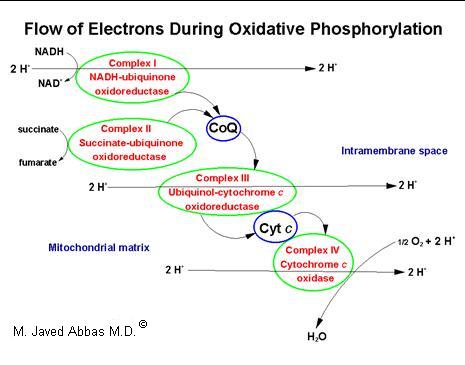
| Diagrammatic representation of the flow of electrons from either NADH or succinate to oxygen (O2) in the electron transport chain of oxidative phosphorylation. Complex I contains FMN and 22-24 iron-sulfur (Fe-S) proteins in 5-7 clusters. Complex II contains FAD and 7-8 Fe-S proteins in 3 clusters and cytochrome b560. Complex III contains cytochrome b, cytochrome c1 and one Fe-S protein. Complex IV contains cytochrome a, cytochrome a3 and 2 copper ions. As electrons pass through the proteins of complex I 4 protons (H+) are pumped into the intramembrane space of the mitochondrion. Two protons are pumped into the intramembrane space as electrons flow through complexes II, III and IV. These protons are returned to the matrix of the mitochondrion, down their concentration gradient, by passing through ATP synthase coupling electron flow and proton pumping to ATP synthesis. |