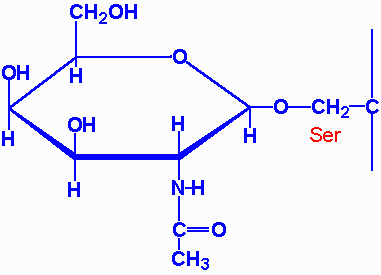
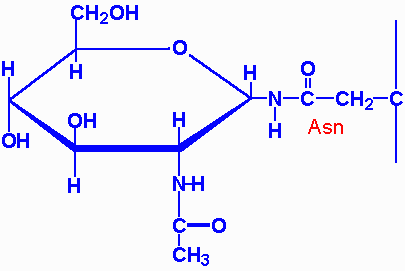
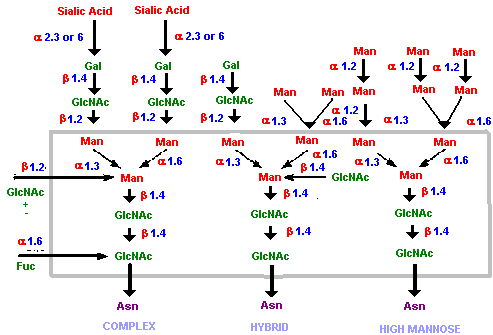
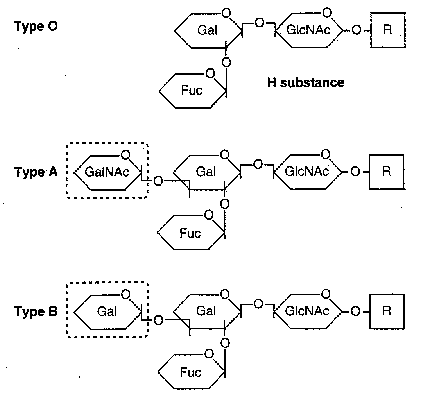
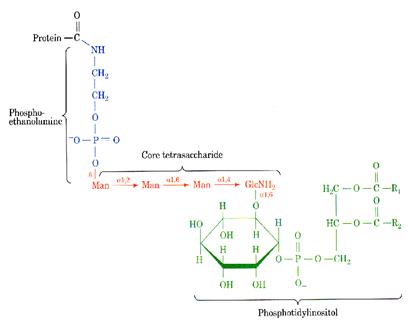
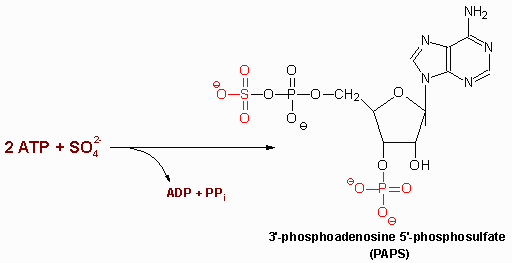
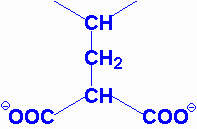
| aspartylglycosaminuria | aspartylglycosaminidase | progressive mental retardation, delayed speech and motor development, coarse facial features
| b-Mannosidosis | b-Mannosidase | primarily neurological defects, speech impairment |
| a-Mannosidosis | a-Mannosidase | mental retardation, dystosis multiplex, hepatosplenomegaly, hearing loss, delayed speech |
| GM1 Gangliosidosis | b-Galactosidase | also identified as a glycosphingolipid storage disease |
GM2 Gangliosidosis
(Sandhoff-Jatzkewitz disease) | b-N-acetylhexosaminidases A and B | also identified as a glycosphingolipid storage disease |
Sialidosis
(also identified as Mucolipidosis I) | Neuraminidase
(sialidase) | myoclonus, congenital ascites, hepatosplenomegaly, coarse facial features, delayed mental and motor development |
| Fucosidosis | a-Fucosidase | progressive motor and mental deterioration, growth retardation, coarse facial features, recurrent sinus and pulmonary infections |
|