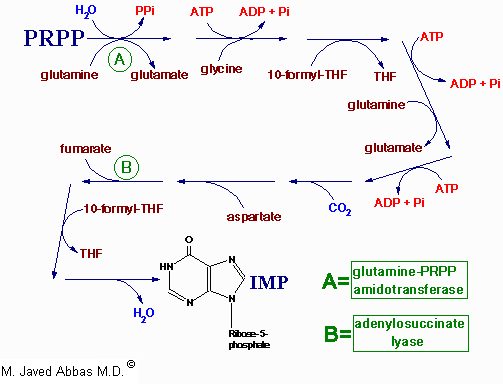
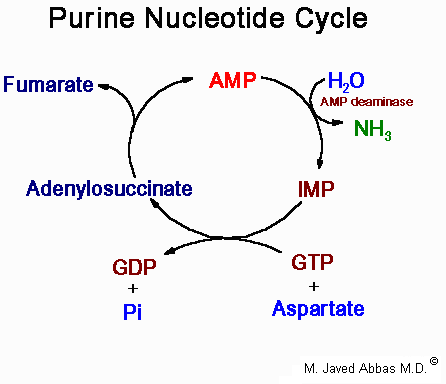
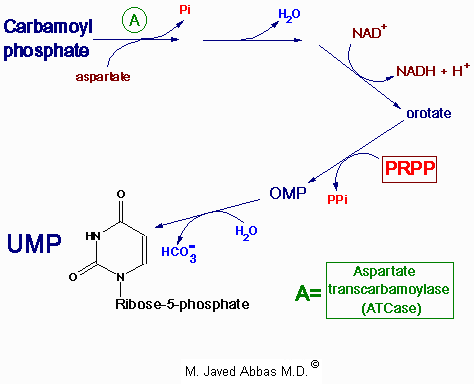
| Disorder | Defective Enzyme | Comments |
| Orotic aciduria, Type I | orotate phosphoribosyl transferase and OMP decarboxylase | see above |
| Orotic aciduria, Type II | OMP decarboxylase | see above |
| Orotic aciduria (mild, no hematologic component) | the urea cycle enzyme, ornithine transcarbamoylase, is deficient | increased mitochondrial carbamoyl phosphate exits and augments pyrimidine biosynthesis; hepatic encephalopathy |
| b-aminoisobutyric aciduria | Transaminase, affects urea cycle function during deamination of a-amino acids to of a-keto acids | benign, frequent in Orientals |
| Drug induced orotic aciduria | OMP decarboxylase | Allopurinol and 6-azauridine treatments cause orotic acidurias without a hematologic component; their catabolic by-products inhibit OMP decarboxylase |