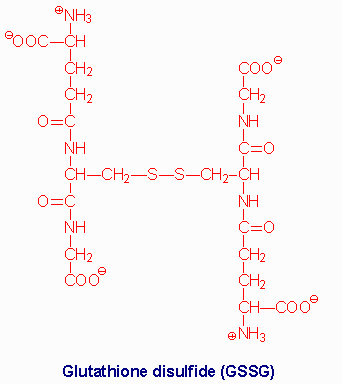| Glutathione (GSH) is a tripeptide composed of
g-glutamate, cysteine and glycine. The sulfhydryl
side chains of the cysteine residues of two glutathione molecules form a
disulfide bond (GSSG) during the course of being oxidized in reactions
with various oxides and peroxides in cells. Reduction of GSSG to two moles
of GSH is the function of glutathione reductase, an enzyme
that requires coupled oxidation of NADPH.
|