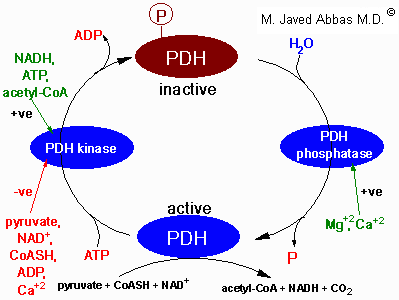
| Factors regulating the activity of
pyruvate dehydrogenase, (PDH). PDH activity is regulated by
its' state of phosphorylation, being most active in the dephosphorylated
state. Phosphorylation of PDH is catalyzed by a specific PDH
kinase. The activity of the kinase is enhanced when cellular
energy charge is high which is reflected by an increase in the level of
ATP, NADH and acetyl-CoA. Conversely, an increase in pyruvate strongly
inhibits PDH kinase. Additional negative effectors of PDH kinase are ADP,
NAD+ and CoASH, the levels of which increase when energy levels
fall. The regulation of PDH phosphatase is not completely
understood but it is known that Mg2+ and Ca2+
activate the enzyme. In adipose tissue insulin increases PDH activity and in cardiac muscle
PDH activity is increased by catecholamines.
|