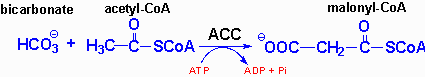
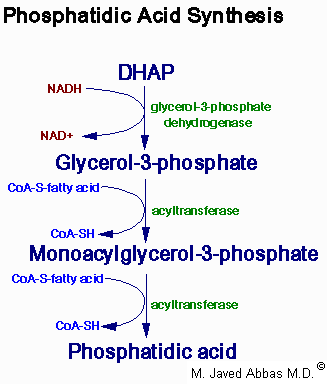
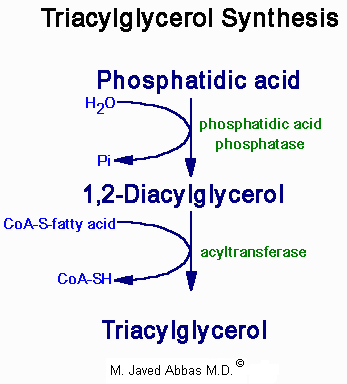
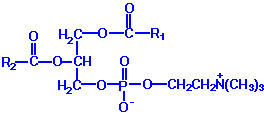
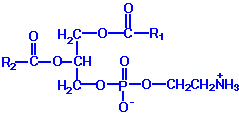
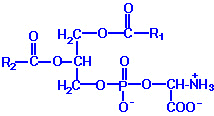
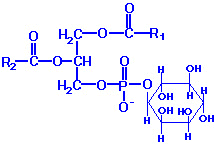
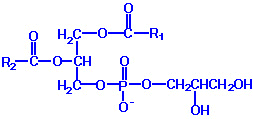
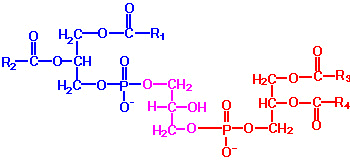
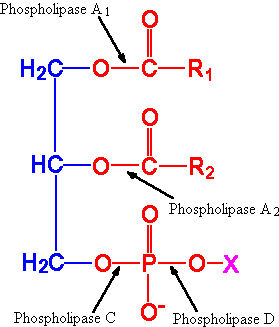
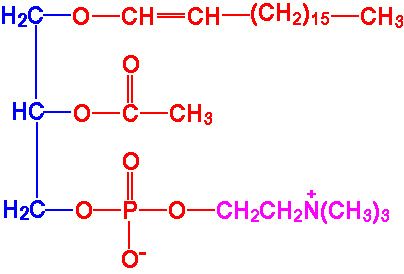
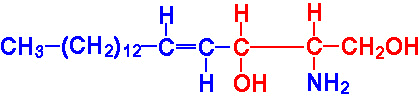
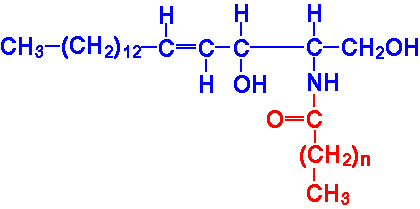
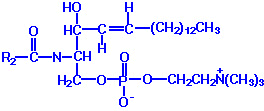
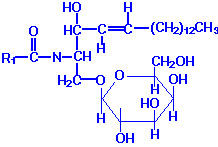
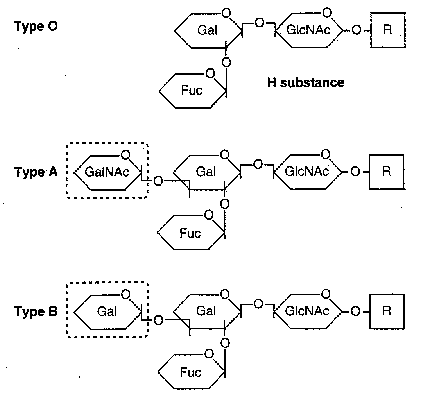
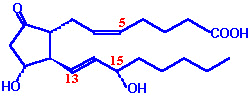
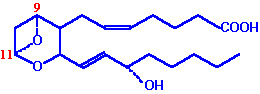
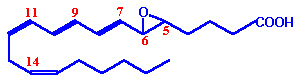
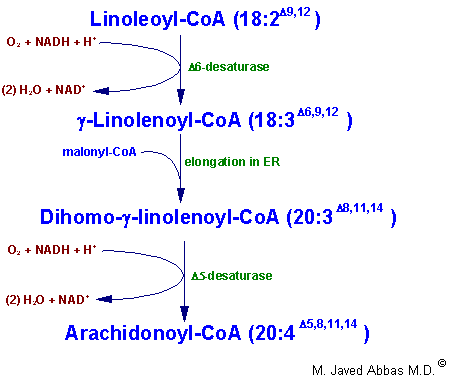
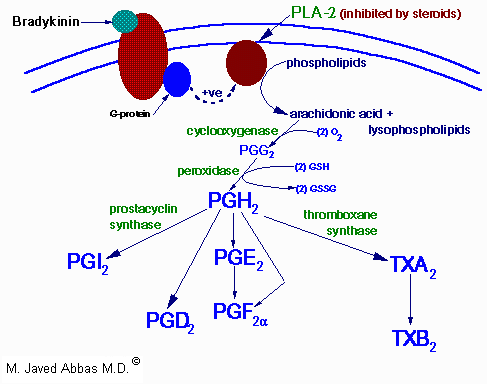
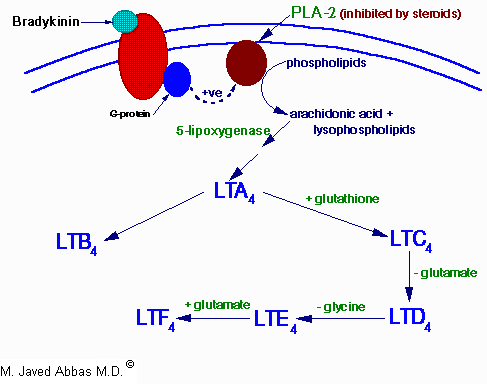
| Eicosanoid | Major site(s) of synthesis | Major biological activities |
| PGD2 | mast cells | inhibits platelet and leukocyte aggregation, decreases T-cell proliferation and lymphocyte migration and secretion of IL-1a and IL-2; induces vasodilation and production of cAMP |
| PGE2 | kidney, spleen, heart | increases vasodilation and cAMP production, enhancement of the effects of bradykinin and histamine, induction of uterine contractions and of platelet aggregation, maintaining the open passageway of the fetal ductus arteriosus; decreases T-cell proliferation and lymphocyte migration and secretion of IL-1a and IL-2 |
| PGF2a | kidney, spleen, heart | increases vasoconstriction, bronchoconstriction and smooth muscle contraction |
| PGH2 | | precursor to thromboxanes A2 and B2, induction of platelet aggregation and vasoconstriction |
| PGI2 | heart, vascular endothelial cells | inhibits platelet and leukocyte aggregation, decreases T-cell proliferation and lymphocyte migration and secretion of IL-1a and IL-2; induces vasodilation and production of cAMP |
| TXA2 | platelets | induces platelet aggregation, vasoconstriction, lymphocyte proliferation and bronchoconstriction |
| TXB2 | platelets | induces vasoconstriction |
| LTB4 | monocytes, basophils, neutrophils, eosinophils, mast cells, epithelial cells | induces leukocyte chemotaxis and aggregation, vascular permeability, T-cell proliferation and secretion of INF-g, IL-1 and IL-2 |
| LTC4 | monocytes and alveolar macrophages, basophils, eosinophils, mast cells, epithelial cells | component of SRS-A, induces vasodilation, vascular permeability and bronchoconstriction and secretion of INF-g |
| LTD4 | monocytes and alveolar macrophages, eosinophils, mast cells, epithelial cells | predominant component of SRS-A, induces vasodilation, vascular permeability and bronchoconstriction and secretion of INF-g |
| LTE4 | mast cells and basophils | component of SRS-A, induces vasodilation and bronchoconstriction |