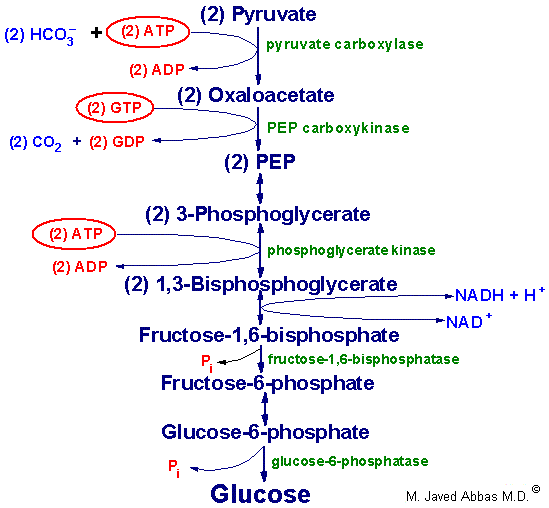
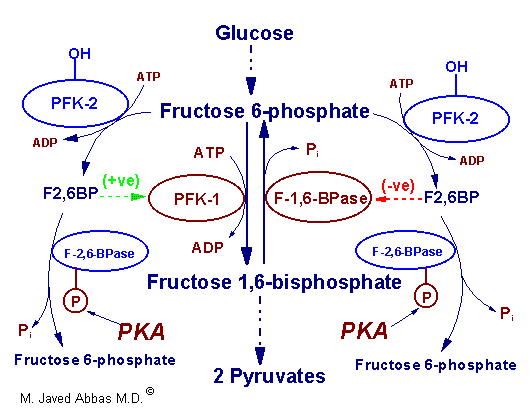
| Regulation of glycolysis and
gluconeogenesis by fructose 2,6-bisphosphate
(F2,6BP). The major sites for regulation of glycolysis and
gluconeogenesis are the phosphofructokinase-1 (PFK-1) and
fructose-1,6-bisphosphatase (F-1,6-BPase) catalyzed
reactions. PFK-2 is the kinase activity and F-2,6-BPase is the phosphatase
activity of the bi-functional regulatory enzyme,
phosphofructokinase-2/fructose-2,6-bisphosphatase. PKA is cAMP-dependent
protein kinase which phosphorylates PFK-2/F-2,6-BPase turning on the
phosphatase activity. (+ve) and (-ve) refer to positive and negative
activities, respectively. |